2018-07-20
前言
今年5月,David R. Liu教授与张锋教授等人联合创立Beam Therapeutics公司,再一次将“单碱基编辑技术”送上了热搜榜,趁着热度还未散去,我们继上一期“单碱基编辑技术浅谈”后,又为大家准备了一篇超级实用性的深度分析,带你一起看看单碱基编辑的前世今生及如何一步步发展壮大的历程!希望大家看后能对单碱基编辑技术有一个更全面的认识。
单碱基基因编辑技术(base editors,BEs)
单碱基基因编辑技术,顾名思义,指能在基因组上引起单个碱基改变的基因编辑技术。基本原理是将胞嘧啶脱氨酶(APOBEC)或腺苷脱氨酶与现存Cas9n(D10A)融合而形成,依赖于CRISPR原理使得靶点远离PAM端的4-7位的单个碱基发生修改的基因编辑技术。
目前的单碱基基因编辑包括两种:
1. CBEs(Cytidine base editors)(图-1)[1]--嘧啶碱基转换技术(C/G到T/A):
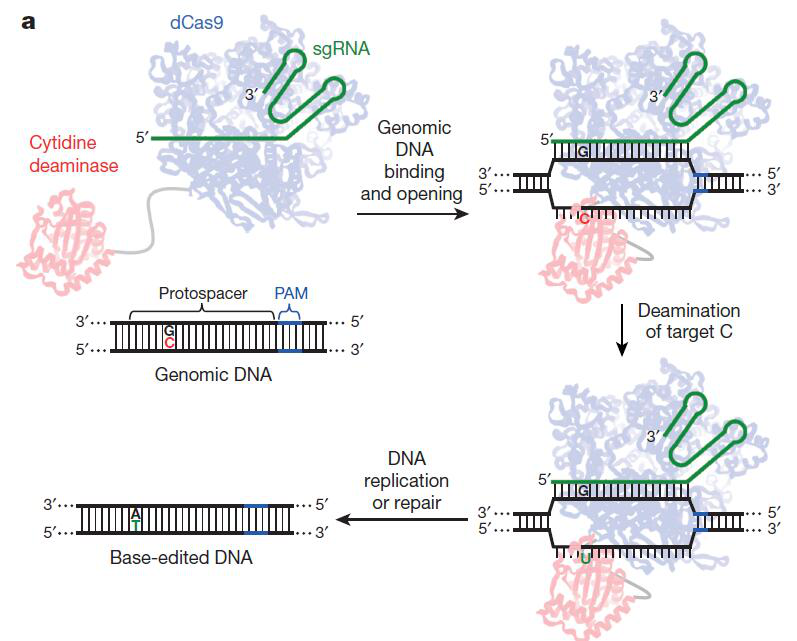
图-1 CBEs工作示意图[1]
2. ABEs(Adenine base editors)(图-2)[2]-嘌呤碱基转换技术(A/T到G/C):
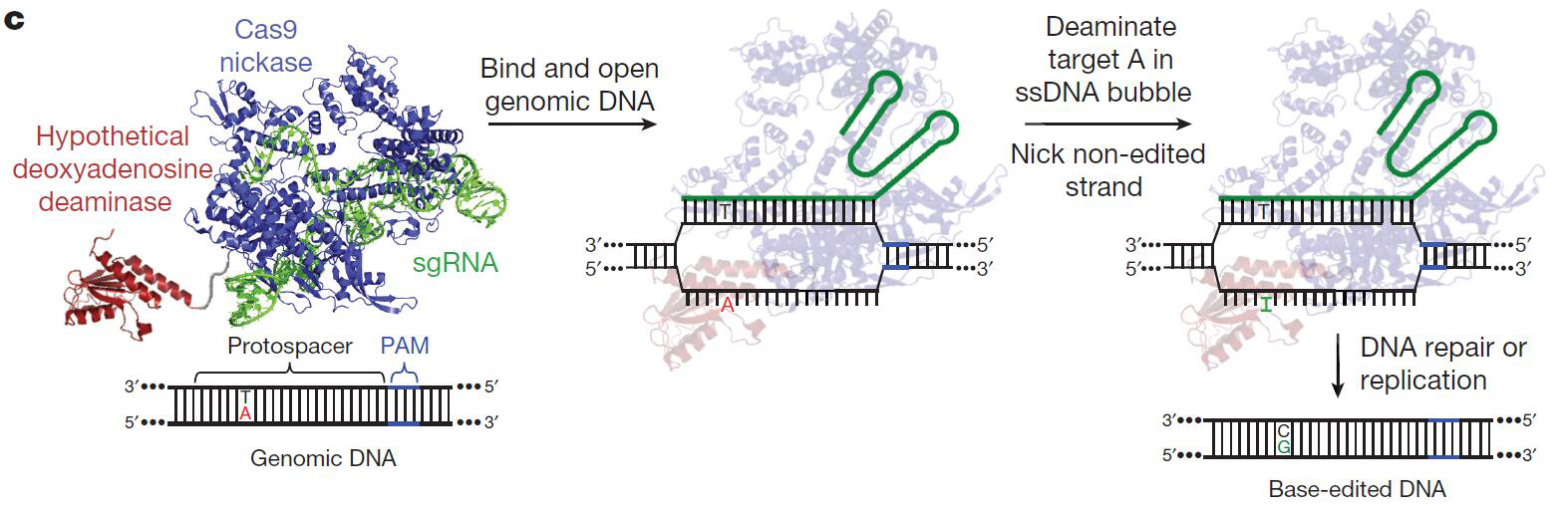
图-2 ABEs工作示意图[2]
相对于传统的CRISPR/Cas9的优势:
传统的CRISPR/Cas9
单碱基编辑技术
编辑效率
细胞:0.1-5%; 胚胎:70%-80%
细胞:70-80%; 胚胎:100%
插入与缺失突变
较高
较低
脱靶效应
较高
较低
单碱基编辑技术工具的发展及优化历程:
单碱基基因编辑技术自2016年4月被报道以来,迅速成为基因编辑技术领域的一颗璀璨的“明星”。其技术的发展核心主要是工具的不断优化。下面具体介绍:
第一类:嘧啶碱基转换工具-- CBE (Cytidine base editor)的优化历程
l 2016年4月: David Liu 实验室首先报道了基于来源于大鼠胞嘧啶脱氨酶与CRISPR/Cas9 融合形成的单碱基基因编辑工具(Cytidine base editors, CBEs),包括BE1, BE2,BE3。其中在他们实验室报道的BE3(图-3),效率最高,也是目前应用的最为广泛的嘧啶碱基编辑工具,目前BE3已应用到基因编辑编辑,基因治疗,动物模型制作及功能基因筛选等领域。

图3. David Liu 实验室报道的BE3工具
l 2016年 8月:日本科学家 Keiji Nishida 等将来源于七鳃鳗的胞嘧啶脱氨酶与 CRISPR/Cas9和尿嘧啶糖苷酶抑制剂融合,该系统可在哺乳动物细胞中实现 15%-55%靶向突变[3]。
l 2016年 10月:常兴课题组将人源胞嘧啶脱氨酶融合 dCas9 的 C 端 (dCas9-AIDx)形成的TAM(targeted AID-mediated mutagenesis)可通过同样可在远离PAM的5-9位产生C到T,A,G的突变[4]。
l 2016年 11月:报道的 CRISPR-X(图-4),将来源于人的胞嘧啶脱氨酶突变体去除出核信号后融合到MS2蛋白上,通过 MS2 RNA 的发夹结构招募到sgRNA 的骨架中,经优化后的CRISPR-X, 可以引起+20到+40bp 窗口内实现平均20.6%的突变效率[5]。
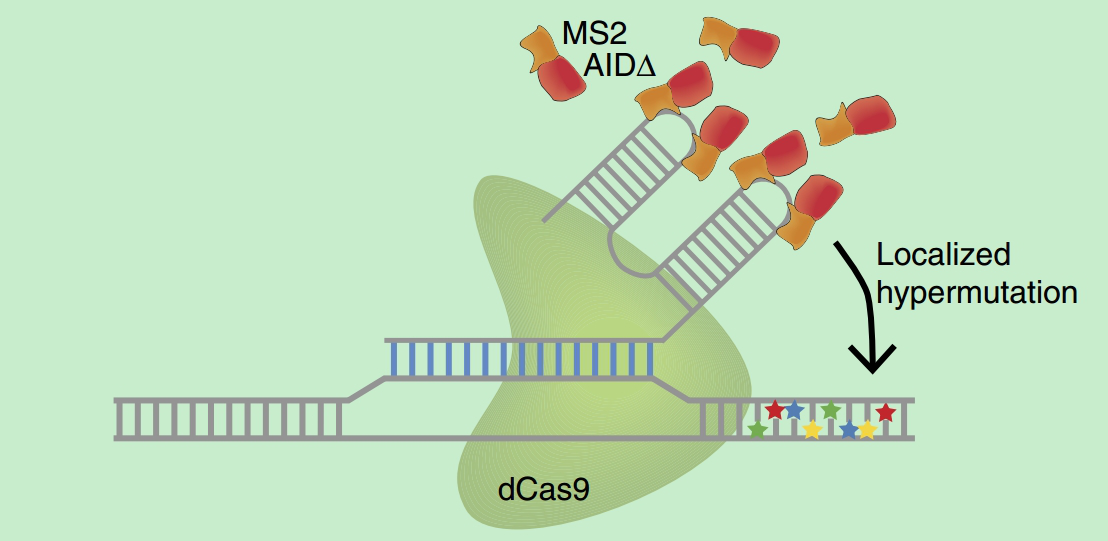
图4. CRISPR-X工作原理图[5]
l 2016年 11月:杨璐菡团队等将TALE, Zinc-finger DNA特异识别单元与来源于人的胞嘧啶脱氨酶进行融合,可在细菌和人的细胞中分别实现 13% 和 2.5%的 C 到 T 的突变效率[6],这可能是由于胞嘧啶脱氨酶的底物是单链DNA, 而 TALE和 Zinc-finger 无法打开双链 DNA从无法使单链DNA暴露于胞嘧啶脱氨酶,影响了脱氨酶的效率,进而影响了C 到 T 的转换效率。
l 2017年1 月: 同时David Liu 实验室也在积极探索不受PAM限制的新型高效的单碱基因组编辑工具, 如VQR-BE3(PAM: NGAN), SaKKH-BE3(NNNRRT)等。 同时,他们也通过胞嘧啶脱氨酶功能域进行氨基酸突变进而筛选到编辑窗口能精确到1-2碱基的单碱基工具,其中以YE1-BE3(W90Y+R126E)为最优,保持与BE3相似编辑活性的同时,将编辑窗口精确到1-2个碱基[7]。
l 2018年5 月:上海科技大学陈佳实验室将Cpf1与胞嘧啶脱氨酶进融合,由于Cpf1特殊结构,无法设计成切口靶向链而将非靶向链暴露于脱氨酶,因此,最初的Cpf1-Apobec1效率并不高,这一点与TALE-AID类似。但通过将核定位信号引入Cpf1-Apobec1改造为dCpf1-BEs (图-5)使其工作效率最高可达40%以上[8],因此,表明NLS的引入在单碱基基因编辑中有着至关重要的作用。
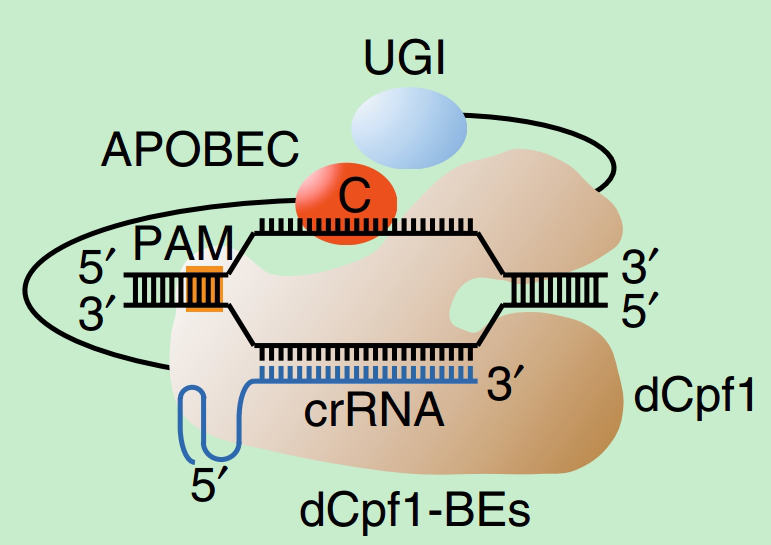
图-5 dCpf1-BEs工作原理图[8]
l 通过扩充PAM的CBE,将使CBE工具的使用更加灵活。 但由于CBE固有的原理,即胞嘧啶发生脱氨基时,随后将刺激细胞内的切除修复,因而在碱基编辑过程中不可避免的产生较低频的indels及非预期突变(C到G,A的突变),降低了其产物纯度。 鉴于此,2017年8月, David Liu 实验室及陈佳实验室通过优化linker 及UGI的表达量,通过抑制内源糖苷酶活性,得到BE4 和eBE-S3(图-6)均可以不同程度地降低indels及非预期突变,提高其产物纯度[9, 10]。


图-6. BE4 和eBE-S3
因引,经过科学家们对CBE工基础上进行了深入的挖掘和优化,目前的CBE已可以做到较高的编辑效率,较高的产物纯度,较精确的编辑窗口,较灵活PAM选择,这在精确碱基替换方面展示了无可替代的优势。
第二类:嘌呤碱基转换工具--ABEs(Adenine base editors)的优化历程
对于2017年10月报道的嘌呤转换工具-ABEs,它打破了过去单碱基编辑仅能编辑嘧啶碱基的限制。 仅仅过去半年之多, 其已经在基因编辑,动物模型制作,基因治疗,植物的基因修饰等领域相续报道。 它的原理与CBE稍有不同,即腺嘌呤脱氨基之后为肌苷,在DNA水平被当作G进行读码复制,然而细胞内对于肌苷的切除修复并不敏感,因此,ABEs能够较为高效的产生A/T到G/C的突变,同时维持较高的产物纯度。
l 2018年2月: David Liu 实验室,考虑到仅有PAM仍然是其使用的一个限制,后来David Liu实验室将Cas9通过PACE进化得到了不同氨基酸突变的xCas9 (图-7)其中以xCas9 3.7最优, 它不仅NGG PAM与传统的Cas9保持相当的活性,而且可以,识别 NGA,NGC,NGT, GAA,GAT, 同时保持较低的脱靶效应,这样可以靶向人类基因组范围内约的1/16的靶点。 之后将其与ABEs 融合,也同样使ABEs PAM的选择也更加灵活[11]。
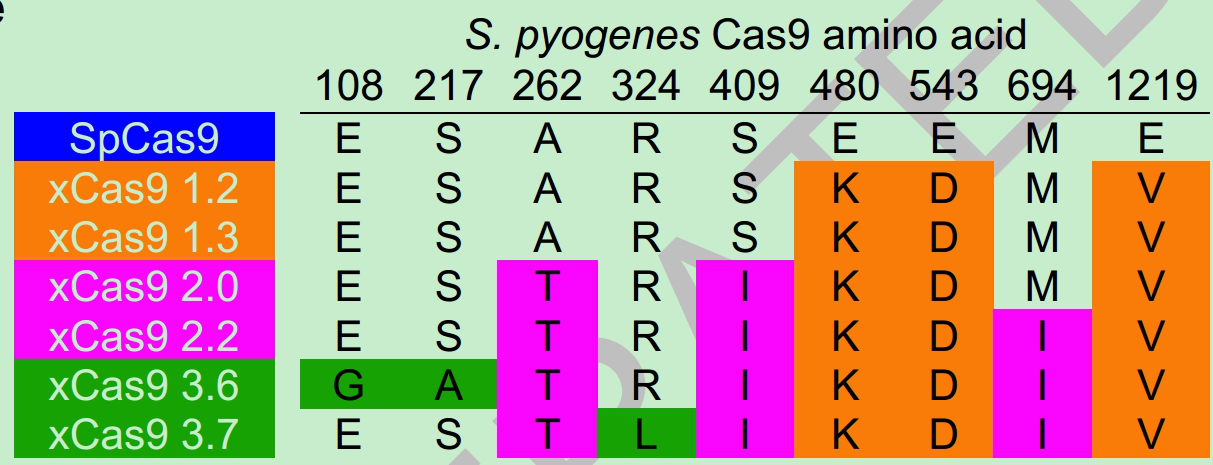
图7. 不同氨基酸突变的xCas9(来自网络)
展望ABEs的发展,就是回首看CBE的发展。因此,对于ABEs来讲,更加灵活的PAM工具的开发,如 SaKKH(NNNRRT),Cpf1(TTTN) 的融合,也显得尤为必要。
l 2018年5月: 另外,最近David Liu实验室,在nature biotechnology报道通过密码子优化后的序列以及引入额外的核定位信号(双分型)的base editors, 得到BE4max 和ABEmax可以分别提高1.9倍,1.3-7.9倍的碱基编辑效率[13]。
l 2018年 7月:报道了BE-Plus (图-8),即将多个多肽GCN4与其抗体结合区分别融合Cas9n(D10A),Apobec1-UGI, 它可以募集多个拷贝的Apobec1-UGI的融合蛋白,这样可以在靶点区3-16位 C/G 到T/C的转变,可实现较大范围内单个碱基突变[12]。类比,将其运用到ABEs当中,也同样可以实现较大范围内A/T到G/C的突变。
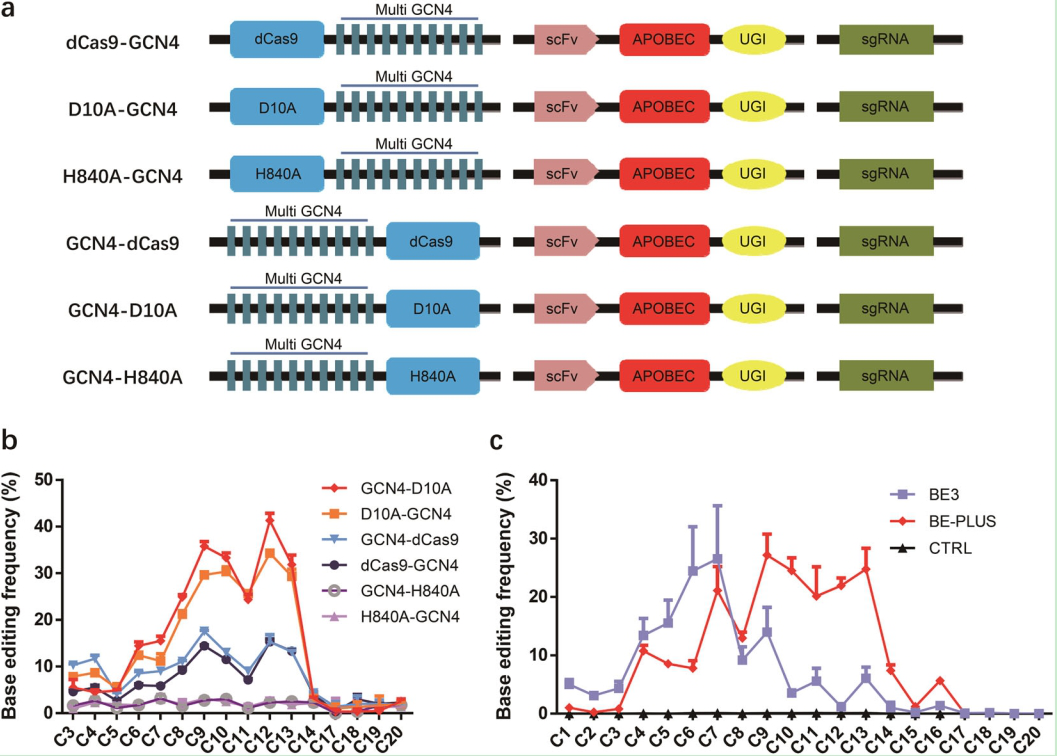
图8. BE-Plus的编辑窗口及编辑效率
l 2018年7月3日:来自nature biotechnology上最新报道, 也同样将密码子优化序列(图-9-1)及核定位序列引入(图-9-2)到单碱基中BE3中,以FNLS为例(图-9-3),用慢病毒在较为难转染的细胞中(如NIH3T3细胞)最高提高了约50倍的C到T 的突变(相对于BE3)(图-9-2),使应用更加广泛[14]。这些优化将促使单碱基基因编辑工具更加高效,灵活,通用。

图-9-1:RA:Lenti-BE3 密码子优化
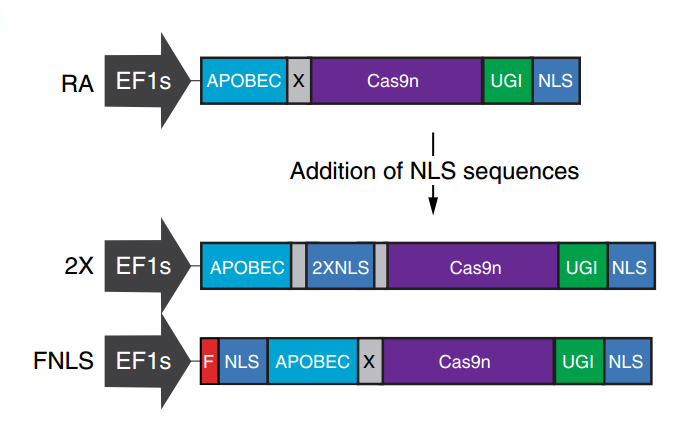
图-9-2 FNLS:RA的N端引入核定位序列
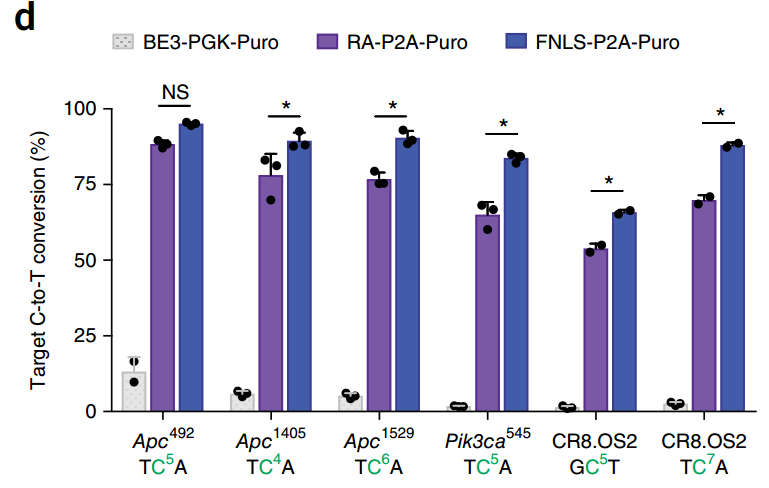
图-9-3 优化后不同Lenti-BE3 在NIH3T3细胞中不同的靶点编辑效率
单碱基编辑技术的应用
点突变是引起人类遗传病的主要原因之一。据 ClinVar database显示,C/G 到 T/A, A/T 到 G/C突变引起致病单核苷酸突变的分别占48%,14% (图-10)。
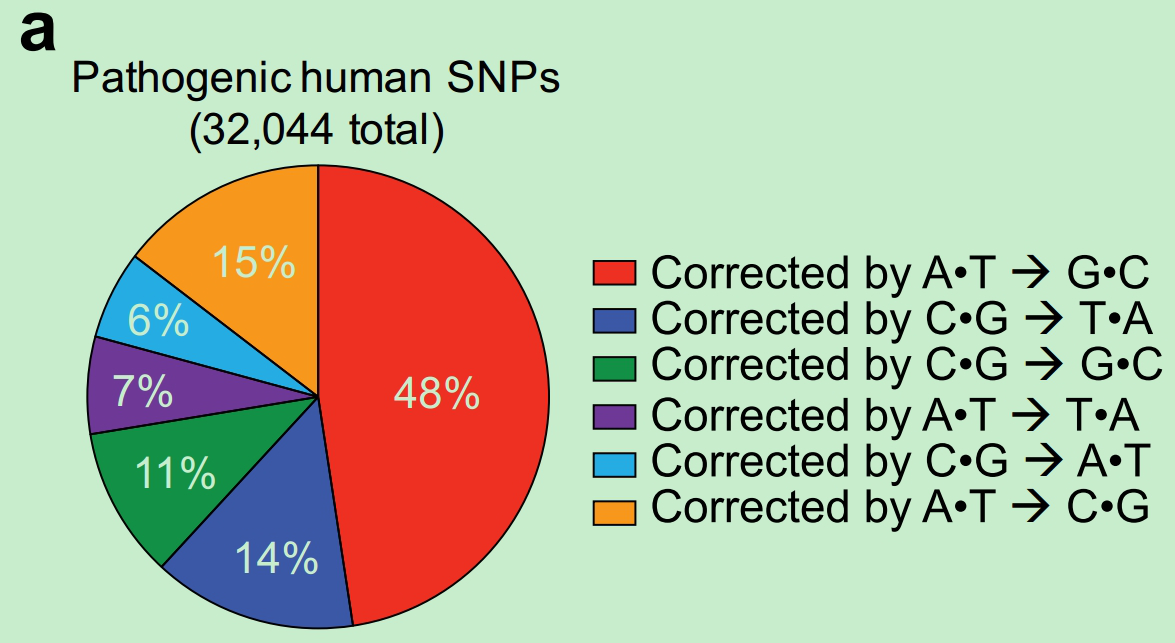
图-10 人类遗传病中单碱基突变类型
也即是,这两种类型的突变将极大可能被ABE或CBE进行较正。因此,未来两种高效灵活的单碱基工具将会在未来的基因治疗中大显身手。如β-血红蛋白病, 它主要由于在成年人中β-globin基因突变导致功能异常所致。 目前已证实 高水平表达胎儿血红蛋白(HbF)病被认为治疗β-血红蛋白病的很有潜力的策略[15]。-198 T > C[16], -175 T > C[17]这些在β-globin的启动区的突变可以通过不同的机制提高 HbF的表达。而这些突变已被报道可以被单碱基工具ABE7.10高效编辑[2, 13]。因此,未来利用单碱基治疗β-血红蛋白病(如地中海贫血)将极有可能是一个绝佳的策略。
虽然,CBE或ABE单碱基编辑技术具有其强大的优势,但是离未来临床疾病治疗仍然还有很多问题需要解决。
1. 主要是以下几点:如CBE,ABE体积都过大,在基因治疗中需要进行有效拆分后包装进AAV进行体内治疗;
2. 典型的CBE,ABE 高效编辑窗口依然是4-7位, 是否可将实现窗口平移,以靶向基因组更为广阔的空间也有待进一步研究;
3. CBE可以通过突变胞嘧啶脱氨酶特定位点从而实现更加精确的1-2位的编辑窗口,而相对ABE的窗口最为精确也只能为4-7位,能否进一步通过突变腺嘌呤脱氨酶特定位点从而实现更加精确的ABE的编辑窗口,有待进一步研究。
4. 在CBE,ABE中虽然均能高效地编辑某些靶点,但并非所有靶点均能编辑,其高效靶点是否存在某种规律?是否也Cas9中靶点的选择存在某种关系,也有待研究。
5. 目前已经实现嘧啶与嘧啶,嘌呤与嘌呤之间互换,是否存在嘧啶与嘌呤之间的碱基互换,也有待进一步研究;
6. 目前,Cas9蛋白结构已知,胞嘧啶脱氨酶(APOBEC)或腺苷脱氨酶的蛋白结构也未知,融合成相应的CBE,ABE结构也未知,因此,未来解析CBE,ABE的融合蛋白结构深入理解单碱基编辑靶向识别与编辑机制,也意义重大。
参考文献:
[1] Komor AC, Kim YB, Packer MS, et al. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature, 2016, 533: 420
[2] Gaudelli NM, Komor AC, Rees HA, et al. Programmable base editing of a• t to g• c in genomic DNA without DNA cleavage. Nature, 2017, 551: 464
[3] Nishida K, Arazoe T, Yachie N, et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science, 2016, 353: aaf8729
[4] Ma Y, Zhang J, Yin W, et al. Targeted aid-mediated mutagenesis (tam) enables efficient genomic diversification in mammalian cells. Nature methods, 2016, 13: 1029
[5] Hess GT, Frésard L, Han K, et al. Directed evolution using dcas9-targeted somatic hypermutation in mammalian cells. Nature methods, 2016, 13: 1036
[6] Yang L, Briggs AW, Chew WL, et al. Engineering and optimising deaminase fusions for genome editing. Nature communications, 2016, 7: 13330
[7] Kim YB, Komor AC, Levy JM, et al. Increasing the genome-targeting scope and precision of base editing with engineered cas9-cytidine deaminase fusions. Nature biotechnology, 2017, 35: 371
[8] Li X, Wang Y, Liu Y, et al. Base editing with a cpf1–cytidine deaminase fusion. Nature biotechnology, 2018, 36: 324
[9] Wang L, Xue W, Yan L, et al. Enhanced base editing by co-expression of free uracil DNA glycosylase inhibitor. Cell research, 2017, 27: 1289
[10] Komor AC, Zhao KT, Packer MS, et al. Improved base excision repair inhibition and bacteriophage mu gam protein yields c: G-to-t: A base editors with higher efficiency and product purity. Science advances, 2017, 3: eaao4774
[11] Hu JH, Miller SM, Geurts MH, et al. Evolved cas9 variants with broad pam compatibility and high DNA specificity. Nature, 2018, 556: 57
[12] Jiang W, Feng S, Huang S, et al. Be-plus: A new base editing tool with broadened editing window and enhanced fidelity. Cell research, 2018, 1
[13] Koblan LW, Doman JL, Wilson C, et al. Improving cytidine and adenine base editors by expression optimization and ancestral reconstruction. Nature biotechnology, 2018,
[14] Zafra M, Schatoff E, Katti A, et al. Optimized base editors enable efficient editing in cells, organoids and mice. Nature biotechnology, 2018,
[15] Traxler EA, Yao Y, Wang Y-D, et al. A genome-editing strategy to treat β-hemoglobinopathies that recapitulates a mutation associated with a benign genetic condition. Nature medicine, 2016, 22: 987
[16] Wienert B, Martyn GE, Kurita R, et al. Klf1 drives the expression of fetal hemoglobin in british hpfh. Blood, 2017, blood-2017-2002-767400
[17] Wienert B, Funnell AP, Norton LJ, et al. Editing the genome to introduce a beneficial naturally occurring mutation associated with increased fetal globin. Nature communications, 2015, 6: 7085

